
Fungi have the following characteristics:
- basic structural unit is almost always a hypha, a branched filament of cells (a, b)
- reproductive propagule almost always a spore (c, usually single-celled)
- nutrition is heterotrophic and absorptive (they secrete enzymes that break down organic matter, then absorb soluble products)
Other important characteristics:
- mostly non-motile
- rigid cell wall usually containing chitin
- indeterminate growth
What role do fungi play in the forest?
- saprobes (consuming and recycling dead organic matter)
- pathogens (causing plant diseases)
- mutualistic symbionts (mycorrhizae, lichens)
You may be asking yourself, “Why do I need to learn about fungi? I want to learn about tree diseases and what to do about them.” Good question. You will find that fungi are overwhelmingly the most important pathogens of trees. In order to understand the diseases they cause, how the disease spreads, etc., you have to understand the fungi themselves to some extent.

If you are to understand diseases, it is important that you work very hard, now, to master the overall classification of the fungi and the related details. Do it now and really make sure you’ve got it. If you wait, you will build your knowledge of forest pathology on a shaky foundation. It’s up to you.
You should know well the phyla and most of the subgroups.
Major Groups of Fungi
Phyla, subgroups, and some major genera of fungi important in tree diseases.| Phylum | Subgroup | Some major genera |
|---|---|---|
| Oomycota | Oomycetes | Phytophthora, Pythium (water molds and downy mildews) |
| Glomeromycota | Glomeromycetes | Acaulospora, Gigaspora, Glomus (arbuscular/endomycorrhizal fungi) |
| Zygomycota | Endogonales | Endogone (ectomycorrhizal fungi, saprobes) |
| Mucorales | Mucor, Rhizopus (mostly common saprobes) | |
| Ascomycota | Classification of the Ascomycota is so complex, it is better for the beginning student to focus instead on forms of fruiting as presented here | |
| naked asci | Taphrina | |
| perithecia | Nectria, Ceratocystis, Ophiostoma, Valsa | |
| cleistothecia | Microsphaera | |
| apothecia | Peziza, Rhizina, Rhytisma (saprobes, root pathogens, ectomycorrhizal fungi) | |
| pseudothecia | Apiosporina, Phaeocryptopus | |
| Asexual forms of Ascomycota | ||
| simple conidiophores | Verticillium Penicillium |
|
| coremium | Graphium | |
| sporodochium | Fusarium | |
| acervulus | Gloeosporium Marssonina |
|
| pycnidium | Sphaeropsis Cytospora |
|
| Basidiomycota | Agaricales | Mushrooms (Armillaria), puffballs and bird's nest fungi (Lycoperdon, Crucibulum), and coral fungi (Clavaria) |
| Boletales | Poroid mushrooms (Boletus), gasteroid fungi (Pisolithus, Scleroderma) and merulioid crust fungi (Coniophora, Serpula) | |
| Corticiales | A small residual group of mostly corticioid fungi; not important in forest pathology (Corticium, Vuilleminia) | |
| Hymenochaetales | Polypores (Inonotus, Onnia, Oxyporus, Phellinus, Trichaptum) and crust fungi (Hymenochaete, Hyphoderma, Hyphodontia) | |
| Polyporales | Polypores (Fomitopsis, Ganoderma, Oligoporus, Phaeolus, Polyporus) and corticioid fungi (Phanerochaete, Pulcherricium) | |
| Russulales | Polypores (Albatrellus, Bondarzewia, Heterobasidion), tooth fungi (Echinodontium, Hericium), crust/stereoid fungi (Peniophora, Scytinostroma, Stereum) and mushrooms (Lactarius, Russula) | |
| Pucciniomycetes | Cronartium (rusts), Helicobasidium | |
Above includes mostly groups with important tree pathogens and others that forest pathologists should be at least minimally familiar with. This classification is based on a new higher-level phylogenetic classification of the fungi. Fungi are less well understood than many other groups of organisms and their relationships are just beginning to come into focus, so there are many different classifications in various books. Unfortunately, in many cases, morphology does not correspond with phylogenetic relationships based on molecular data. For an embryonic attempt at classifying all organisms on the internet, see the Tree of Life.
Oomycota
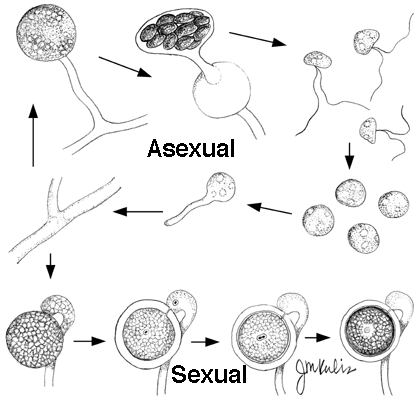
All the groups listed above are in the kingdom Fungi except this one. Depending on one’s kingdom-level classification, this phylum may be placed in kingdom Chromista, Straminipila (=Stramenopila), Protoctista, or Protista.
Your professor may tell you that these are not fungi. While it is true that they are not in the kingdom Fungi, they can legitimately be called fungi (with a small “f”). Think of it as a term that indicates a life form rather than a phylogenetic group, much like ‘trees’. If it looks like a duck, and quacks like a duck, let’s call it a duck.
By the way, this view is supported by the renaming of the relevant code of nomenclature, in 2011, to “International Code of Nomenclature for algae, fungi, and plants”. As noted by Hawksworth [1]:
The lower case letters used for the words “algae”, “fungi”, and “plants” are employed to make clear these terms are being used in a colloquial sense, for instance the inclusion of cyanobacteria in algae, and chromistan fungal analogues and slime moulds in “fungi”.
Unlike true Fungi, Oomycetes (the class of interest in this phylum) have these characteristics:
- sexual spores are oospores formed from unequal gametangia: antheridia transfer nuclei to fertilize oospheres in an oogonium;
- somatic mycelium in a diploid nuclear state;
- cell wall containing cellulose;
- asexual zoospores with two types of flagella (one whiplash, directed posteriorly, the other fibrous, ciliated, directed anteriorly). Some also have nonmotile chlamydospores.
- mitochondria with tubular cristae instead of flattened cristae.


With about 800 species, many important plant pathogens are in this group. Sometimes called “water molds.”
Ascomycota
Sexual spores (ascospores) form in a sac called an ascus (usually in eights), and are often discharged forcibly. The group is often called “ascomycetes” informally.
The drawing at right shows a fully developed ascus containing eight ascospores.
Beside that is an animated ascus developing and shooting out its spores! The complementary, haploid nuclei first undergo mitosis. One pair goes off, perhaps to initiate more asci. The pair that remains is walled off in the ascus initial. They fuse (karyogamy), then undergo meiosis, resulting in four nuceli, then mitosis, to give nuclei for 8 spores. When fully developed, the spores usually shoot out of the ascus under pressure. Complicated little thing, huh?

Naked asci
Sometimes called Hemiascomycetes, now classified in the subphylum Taphrinomycotina. Ascoma absent. These are morphologically simple ascomycetes. Various species of Taphrina cause localized abnormal growth of plants, and asci are produced on the plant surface (right).

Perithecia
Often called Pyrenomycetes, now most are in the class Sordariomycetes. Asci are in a flask-shaped ascoma (perithecium) with a pore (ostiole) at the top (right). This is a big group with some important pathogens.

Cleistothecia
The ascoma (a cleistothecium, right; sometimes called a chasmothecium) is spherical and closed. These are distributed in various modern classes, including Eurotiomycetes, Sordariomycetes and Leotiomycetes.

Apothecia
Often called Discomycetes, most are now in the class Pezizomycetes. The asci are in a bowl- or cup-shaped ascoma (apothecium; right). These are sometimes called the “cup fungi”, and it is a large group.

Pseudothecia
Often called Loculoascomycetes, now placed in the class Dothideomycetes. Asci with two layers (bitunicate), produced in pseudothecia (right) that look like perithecia. Pseudothecia are cavities (locules) in a usually black stroma.

The figure at left shows some asexually reproducing forms of ascomycetes (a, see asexual forms, below), a cleistothecium formed by some powdery mildew fungi (b), apothecia (c), and a vertical section of a perithecium (d).
| Some asexual forms of Ascomycota produce conidia on simple conidiophores with no conidioma (right). |  |
| Conidiophores may also be grouped into columnar coremia (not shown), or cushion-shaped sporodochia (right). |  |
| Conidia may also be produced in an organized, covered fruitbody (conidioma). In one group they are pycnidia (right). They look superficially like perithecia, but of course have no asci. |  |
| In another they are acervuli. |  |
Asexual forms of Ascomycota
Many ascomycetes produce asexual spores (conidia), and often we find the fungus in this state without the sexual form. Some apparently have no sexual reproduction at all. They were traditionally called “deuteromycetes” informally, and were classified in form-taxa from phyla down to species. Such fungi typically had two names, one for the sexual stage which was often used to apply to the “whole fungus”, and one for the asexual stage.
All that changed in 2011 with revisions to the International Code of Nomenclature for algae, fungi, and plants [1]. Dual nomenclature was officially ended. Instead, “One Fungus, One Name” became the rule. Any legitimate name, typified by any stage, can serve as a name for a species, although certain aspects of the rules naturally favored names typified by the sexual stage. As mycologists work to reorganize the chaos of fungal nomenclature, you can expect that there will be related name changes for many years to come! See more details.
Basidiomycota
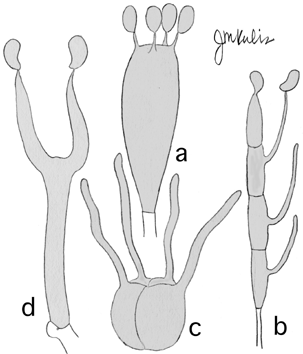
This group is characterized primarily by the sexual spores (basidiosporesbasidium The basidium (pl. basidia) is a cell on which the sexual spores (basidiospores) of basidiomycetes are produced. Usually it is club-shaped, there are four basidiospores on pegs called sterigmata (sing. sterigma), and the spores are forcibly popped off a short distance. In some groups (informally called gasteromycetes), the spores do not pop off.) being produced on a cell called a basidiumbasidium The basidium (pl. basidia) is a cell on which the sexual spores (basidiospores) of basidiomycetes are produced. Usually it is club-shaped, there are four basidiospores on pegs called sterigmata (sing. sterigma), and the spores are forcibly popped off a short distance. In some groups (informally called gasteromycetes), the spores do not pop off., usually in fours. The typical basidium, sometimes called a holobasidium, is aseptate, club-shaped and usually produces four spores (a). Some basidiabasidium The basidium (pl. basidia) is a cell on which the sexual spores (basidiospores) of basidiomycetes are produced. Usually it is club-shaped, there are four basidiospores on pegs called sterigmata (sing. sterigma), and the spores are forcibly popped off a short distance. In some groups (informally called gasteromycetes), the spores do not pop off., such as those of rust fungi, are transversely divided into 4 cells (b). Some (“tremelloid” basidia) are longitudinally divided into 4 cells (c), and others may be shaped like a tuning fork (d).
Members of the Basidiomycota are often called “basidiomycetes” informally. Many, but not all, have special septal structures called clamp connections during most of the life cycle. No other group of fungi has these!

Agaricomycotina

The fungi in this subphylum, about 20,000 described species, have gone through huge changes in classifications in recent years. Although classification now hopefully reflects evolutionary relationships, often there is little in terms of morphology or ecology to help define the groups. In the Agaricomycotina, morphology certainly does not recapitulate phylogeny!
In orders important in forest pathology (class Agaricomycetes, except for Auriculariales, Sebacinales and some members of Cantharellales), the basidia are aseptate and spores germinate to give only hyphae. Basidia are organized in exposed, defined layers (the hymenium) or in an enclosed mass (the so-called gasteroid forms). This class contains fungi that produce the familiar mushrooms, conks, puffballs, stinkhorns, etc.
Of 21 orders currently recognized in the Agaricomycotina, we cover 6 below, all in the class Agaricomycetes. See the outline table above for examples of genera.
Agaricales
More than 9,000 described species, 350 genera and 30 families. Basidioma texture fleshy, basidia usually produced on gills. Most of these are your basic mushrooms, a very large group. A great many agarics are important mycorrhiza partners with trees and shrubs; most of the rest are saprobic decomposers. A few are pathogens. In the bird’s nest fungi (no pathogens), the basidioma is firm and cup-shaped, and fertile tissue is segmented into egg-shaped peridioles: another extravagant and elegant experiment of evolution. Puffballs are modified by production of basidia and spores in a mass inside and enclosed basidioma (gasteroid). There are also some poroid, corticioid, and club/coralloid forms.
Boletales
Most of these are soft, fleshy mushrooms, but with tubes instead of gills (known as boletes). However there are some gasteroid fungi (with basidia and spores in an enclosed mass during development), and some fungi that are flat on the substrate with a wrinkled surface lined with basidia. Most of these fungi are mycorrhizal, but some of the flat ones are important wood decayers, some in living trees and some in wooden buildings.
Corticiales
Mostly corticioid fungi (mostly flat on the substrate with a more or less smooth fertile surface). Traditionally the family Corticiaceae held almost all corticioid fungi and was more important. Now most are distributed in other orders according to their evolutionary relationships with fungi having other forms. No major pathogens remain in this order.
Hymenochaetales
The family Hymonochaetaceae was one of the earliest recognized groups that broke with the traditional classification according to fruitbody form. There are various anatomical and chemical features that clearly unite the members, although the fertile surface in some cases lines tubes (polypores), teeth, and smooth surfaces (corticioid). That family has largely held together under the harsh glare of molecular phylogeny. However, other families, without obvious related features, were formed and added to the order Hymenochaetales. Most members of the order are either polypores or corticioid fungi, and many important pathogens and other wood-decay fungi are found here.
Polyporales

This is still a major group of polypores, including many important pathogens, although other important polypores are found in orders such as Hymenochaetales and Russulales. Some large genera of corticioid fungi are here also. Apparently all members of this order are decayers of wood.
Russulales
Well over 4,000 described species, 80 genera and 12 families. This order now contains just about every configuration of fruitbody. Two very large genera that produce gilled mushrooms form the core of the order, but all the morphological forms found in the other orders are found here as well. Even many of the 12 families within the order have varied forms. Ecologically, they are saprobes, mycorrhizal partners, root parasites and insect symbionts.
Pucciniomycotina and Ustilagomycotina

Basidia usually septate, spores germinate repetitively or by budding. In these two subphyla (rusts and smuts), a basidioma is not formed, karyogamy occurs in a thick-walled resting spore (teliospore), and meiosis occurs upon germination of teliospore.
Pucciniomycotina
Rusts and relatives, more than 8,000 species. Rusts (order Pucciniales, about 7,000 species) are highly specialized parasites of higher plants, life cycles typically with up to five spore stages and two alternating hosts (except by simplification). The rusts cause many serious diseases of economically important hosts, including trees. They also have perhaps the most complex and perplexing life cycles, and variations on those cycles, of any group of organisms. What fun for students!
Ustilaginomycotina
Smuts and relatives, about 1,500 species. Before mating: yeastlike, culturable, usually not infective. After mating: usually obligately parasitic. Many smut fungi have interesting disease cycles that are nicely adapted for plant infection. Infections are often systemic. None are important forest pathogens.
Life Cycle
The life cycles of fungi vary, depending on the group. We will focus on the life cycle of the basidiomycetes, using a polypore as an example.
Life cycle of a typical basidiomycete
This life cycle applies to most wood-decay fungi, especially polypores.| Spores are wind disseminated. Each has a haploid nucleus. We designate the nuclear state as "n." The spores germinate if they are lucky enough to land on a suitable substrate and conditions are favorable. |  |
| When two sexually compatible germlings grow nearby, their hyphae fuse. Mixing the cell contents of the two individuals as a step of the sexual cycle is called plasmogamy. Then there are two different kinds of nuclei together, a state we call the dikaryon ("n+n"). Clamp connections are usually formed in the dikaryon. The fungus exists in this state for most of its life. |  |
| When it has had enough to eat and conditions otherwise allow, the fungus produces a fruitbody, the function of which is sexual reproduction and dispersal. In the polypores, we call them conks informally. Formally, we call them basidiomata (singular basidioma). |  |
| In the lower part of the conk are tubes that end in pores on the lower surface. The tubes are lined with basidia. The basidia are busy little cells, for three important events of the life cycle take place there: karyogamy, meiosis, and spore production. |  |
| This is roughly what a tube looks like in cross section (i.e., looking into the pores). Notice that the spores are forcibly discharged from the basidia, but just far enough to reach the middle of the tube. Then they fall out and are carried away in the wind. |  |
References
- 1.Hawksworth D. 2011. A new dawn for the naming of fungi: impacts of decisions made in Melbourne in July 2011 on the future publication and regulation of fungal names. IMA Fungus 2(2):155–62.
- 2.Worrall JJ, ed. 1999. Structure and Dynamics of Fungal Populations. Dordrecht, The Netherlands: Kluwer Academic Press. 348 pp. <http://dx.doi.org/10.1007/978-94-011-4423-0>.